Modeling of Neuronal Activity in the Visual Cortex
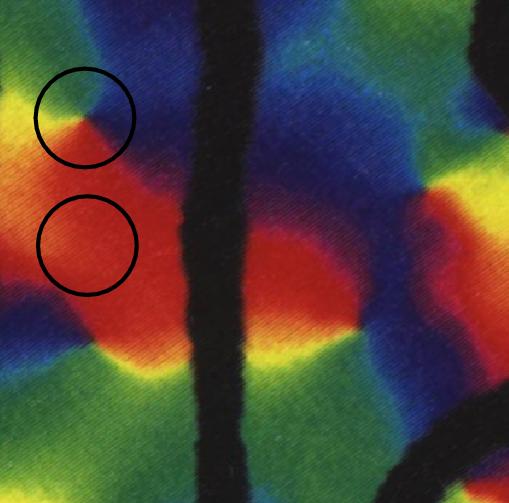
From Blasdel (J. of Neuroscience 12, 1992), the
optical imaging of the orientation hypercolumn structure in monkey visual
cortex.
|

From McLaughlin et al (PNAS 97, 2000), the response
of a model visual cortex to oriented drifting grating stimulus.
|
An effective kinetic representation
of fluctuation-driven neuronal networks with application to simple and complex
cells in visual cortex
D. Cai, L. Tao, M. Shelley, and D. McLaughlin,
in PNAS 101,
7757-7762 (2004).
Abstract: A coarse-grained representation of neuronal network
dynamics is developed in terms of kinetic equations, which are
derived by a moment closure, directly from the original large-scale
integrate-and-fire (I&F) network. This powerful kinetic theory
captures the full dynamic range of neuronal networks, from the
mean-driven limit (a limit such as the number of neurons N

 , in which the fluctuations vanish) to the fluctuation-dominated limit
(such as in small N networks). Comparison with full numerical
simulations of the original I&F network establishes that the
reduced dynamics is very accurate and numerically efficient over
all dynamic ranges. Both analytical insights and scale-up of numerical
representation can be achieved by this kinetic approach. Here,
the theory is illustrated by a study of the dynamical properties
of networks of various architectures, including excitatory and
inhibitory neurons of both simple and complex type, which exhibit
rich dynamic phenomena, such as, transitions to bistability and
hysteresis, even in the presence of large fluctuations. The implication
for possible connections between the structure of the bifurcations
and the behavior of complex cells is discussed. Finally, I&F
networks and kinetic theory are used to discuss orientation selectivity
of complex cells for "ring-model" architectures that characterize
changes in the response of neurons located from near "orientation
pinwheel centers" to far from them.
, in which the fluctuations vanish) to the fluctuation-dominated limit
(such as in small N networks). Comparison with full numerical
simulations of the original I&F network establishes that the
reduced dynamics is very accurate and numerically efficient over
all dynamic ranges. Both analytical insights and scale-up of numerical
representation can be achieved by this kinetic approach. Here,
the theory is illustrated by a study of the dynamical properties
of networks of various architectures, including excitatory and
inhibitory neurons of both simple and complex type, which exhibit
rich dynamic phenomena, such as, transitions to bistability and
hysteresis, even in the presence of large fluctuations. The implication
for possible connections between the structure of the bifurcations
and the behavior of complex cells is discussed. Finally, I&F
networks and kinetic theory are used to discuss orientation selectivity
of complex cells for "ring-model" architectures that characterize
changes in the response of neurons located from near "orientation
pinwheel centers" to far from them.
Download pdf
An Egalitarian Network Model for the Emergence
of Simple and Complex Cells in Visual Cortex
L. Tao, M. Shelley, D. McLaughlin, and R. Shapley,
in PNAS 101, 366-371 (2004).
Abstract: We explain how Simple and Complex cells arise in a
large-scale neuronal network model of the primary visual cortex of the macaque.
Our model consists of approximately 4,000 integrate-and-fire, conductance-based
point neurons, representing the cells in a small, 1 square millimeter patch
of an input layer of the primary visual cortex. In the model the
local connections are isotropic and nonspecific, and convergent input from
the lateral geniculate nucleus confers cortical cells with orientation and
spatial phase preference. The balance between lateral connections
and LGN drive determines whether individual neurons in this recurrent circuit
are Simple or Complex. The model reproduces qualitatively the experimentally
observed distributions of both extracellular and intracellular measures
of Simple and Complex response.
Download pdf
Mexican hats and pinwheels in Visual Cortex
K. Kang, M. Shelley, and H. Sompolinsky,
in PNAS 100, 2848-2853 (2003).
Abstract: Many models of cortical function assume that local
lateral connections are specific with respect to the preferred features of
the interacting cells and that they are organized in a Mexican-hat pattern
with strong center excitation flanked by strong
surround inhibition. However, anatomical data on primary visual
cortex indicate that the local connections are isotropic and that inhibition
has a shorter range than excitation. We address this issue in an analytical
study of a neuronal network model of the local cortical circuit in primary
visual cortex...
Download pdf
States of High Conductance in a Large-Scale Model
of the Visual Cortex
M. Shelley, D. McLaughlin, R. Shapley, and D.J. Wielaard
Journal of Computational Neuroscience 13,
pp. 93-109 (2002).
Abstract: This paper reports on the consequences
of high, activity dependent, synaptic conductances in neurons in a large-scale
neuronal network model of an input layer of the Macaque primary visual
cortex (Area V1)....
Download pdf
Coarse-Grained Reduction and Analysis of a
Network Model of Cortical Response. I. Drifting Grating Stimuli
M. Shelley and D. McLaughlin
Journal of Computational Neuroscience 12, pp.
97-122 (2002)
Abstract: We present
a reduction of a large-scale network model of visual cortex developed
by McLaughlin, Shapley, Shelley, and Wielaard. The reduction is from
many integrate-and-fire neurons to a spatially coarse-grained system for
firing rates of neuronal subpopulations. It accounts explicitly for
``disordered'' properties that vary widely from cortical neuron to cortical
neuron, such as preferred spatial phase. The result is a set of nonlinear
spatio-temporal integral equations for ``phase-averaged'' firing rates across
the model cortex. For drifting grating stimuli, this system yields
time invariant cortico-cortical conductances, with firing rates averaged
over stimulus period being the natural objects. Mathematical analysis
then unveils the mechanisms underlying the spatially varying firing rates
and orientation selectivity observed in the large-scale point-neuron simulations.
This reduction also reproduces, at far less computational cost, the salient
features of the point-neuron network, and is used to study cortical response
to changing stimulus contrast, noise level, and coupling length-scales.
Download pdf
Efficient and Accurate Time-Integration Schemes for Integrate-and-Fire
Neuronal Networks
M. Shelley and L. Tao
Journal of Computational Neuroscience 11, 111-119
(2001).
Abstract: To avoid
the numerical errors associated with resetting the potential following
a spike in simulations of integrate-and-fire neuronal networks, Hansel
et al. and Shelley independently developed a modified time-stepping method.
Their particular scheme consists of second-order Runge-Kutta time-stepping,
a linear interpolant to find spike times, and a recalibration of post-spike
potential using the spike times. Here we show analytically that such a
scheme is second order, discuss the conditions under which efficient, higher-order
algorithms can be constructed to treat resets, and develop a modified
fourth-order scheme. To support our analysis, we simulate a system of
integrate-and-fire conductance-based point neurons with all-to-all coupling.
For 6-digit accuracy, our modified Runge-Kutta fourth-order scheme needs
a time-step of 10^{-3} seconds, whereas to achieve comparable accuracy
using a recalibrated second-order or a first-order algorithm requires
time-steps of 10^{-5} seconds or 10^{-9} seconds, respectively. Furthermore,
since the cortico-cortical conductances in standard integrate-and-fire
neuronal networks do not depend on the value of the membrane potential,
we can attain fourth-order accuracy with computational costs normally associated
with second-order schemes.
Download the PDF
How Simple Cells are made in a Nonlinear Network Model
of the Visual Cortex
D.J. Wielaard, M.J. Shelley, David McLaughlin, and Robert Shapley
Journal of Neuroscience 21, pp. 5203-5211 (2001).
Abstract:
Simple cells in the striate cortex respond to visual stimuli
in an approximately linear manner, even though the LGN input to the striate
cortex, and the cortical network itself, are highly nonlinear...
Download the pdf
A Neuronal Network Model of Macaque Primary Visual Cortex (V1):
Orientation Tuning and Dynamics in the Input Layer 4Calpha.
David McLaughlin, Robert Shapley, Michael Shelley, and D.J. Wielaard
PNAS (2000), v. 97, pp. 8087-8092
Abstract: Here, we offer
an explanation for how selectivity for orientation could be produced by a
model with circuitry that is based on the anatomy of V1 cortex....
Download the PDF
Computational Modeling of Orientation Tuning Dynamics in Monkey Primary
Visual Cortex
M. C. Pugh, D. L. Ringach, R. Shapley and M. J. Shelley
Journal of Computational Neuroscience (2000), v. 8,
pp. 143-159
Abstract: In the primate
visual pathway, orientation tuning of neurons is first observed in the primary
visual cortex. The LGN cells that comprise the thalamic input to V1 are
not orientation tuned, but some V1 neurons are quite selective. Two main
classes of theoretical models have been offered to explain orientation selectivity:
feedforward models, in which inputs from spatially aligned LGN cells are summed
together by one cortical neuron; and feedback models, in which an initial
weak orientation bias due to convergent LGN input is sharpened and amplified
by intracortical feedback. Recent data on the dynamics of orientation tuning,
obtained by a cross-correlation technique, may help to distinguish between
these classes of models. To test this possibility, we simulated the measurement
of orientation tuning dynamics on various receptive field models, including
a simple Hubel-Wiesel type feedforward model: a linear spatio-temporal filter
followed by an integrate-and-fire spike generator. The computational study
reveals that simple feedforward models may account for some aspects of the
experimental data, but fail to explain many salient features of orientation
tuning dynamics in V1 cells. A simple feedback model of interacting cells
is also considered. This model is successful in explaining the appearance
of Mexican-hat orientation profiles, but other features of the data continue
to be unexplained.
click here
for the *.ps.gz file of the article.
click here
for the *.pdf file of the article.